Life Cycle
In the Insect
Vector
The African
trypanosomes reside almost exclusively in the
bloodstream and are transmitted by the bite of the
tse-tse fly which acquires the infection while
taking a bloodmeal, and returns the trypanosome to
a vertebrate host in its saliva when it takes
another bloodmeal. Because this
mode of transmission is by inoculation during
biting this group of trypanosomes are also
referred to as saliva-type or “Salivarian”. (T.
cruzi, on the other hand, is transmitted by
fecal contamination and is referred to as a
“Stercorarian”). The range of African
trypanosomiasis is determined by the range of the
vector. Interestingly, only newly hatched tse-tse
flies are competent to transmit the disease.
Glossina
is in fact a poor vector in nature since less than
1% of the flies are infected.

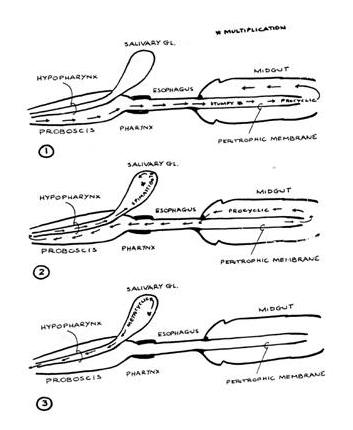
The ingested form
that is infectious for the fly is termed the
short-stumpy bloodstream trypomastigote, which is
a non-dividing form. Following ingestion, the
bloodmeal is retained within the midgut, and the
parasite differentiates into a procyclic form and
divides by binary fission. After about
two weeks some procyclics migrate from the midgut
through the hemocoel eventually reaching the
salivary glands. At this point they differentiate
through an epimastigote stage into a metacyclic
trypomastigote stage, which is a non-dividing form
infectious for the mammalian host. Metacyclic
trypomastigotes are found in the salivary glands ~
20 days after the bloodmeal, and there are ~
40,000 trypomastigotes/bite, but it takes only 400
to initiate an infection.
Click here to see
steaming videos of procyclic T. brucei.
In the
Mammalian Host
The metacyclic
trypomastigotes replicate at the site of
infection. There may be an immune response causing
inflammation (trypanosomal chancre) at the site of
the bite. From there the trypomastigotes move via
the lymphatics to the lymph nodes and then to the
bloodstream. In T. gambiense infection, swollen
cervical (neck) lymph nodes are referred to as Winterbottom’s sign. Long- slender
bloodstream trypomastigotes divide by binary
fission in the bloodstream, generating, on
occasion, short-stumpy forms to continue the cycle
in the tse-tse fly. The long-slender
trypomastigotes are not infectious for the fly.
|

Winterbottom’s sign and needle aspirate of lymph
node |
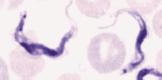
T. brucei trypomastigotes in blood |
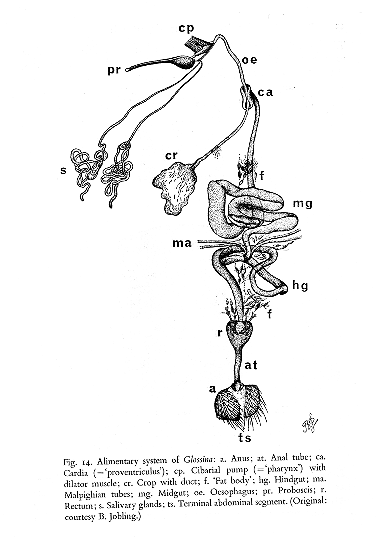
GLUCOSE METABOLISM IN AFRICAN TRYPANOSOMES
The metabolism of the
procyclic trypanosomes in the fly midgut or in culture differs dramatically from
that of the bloodstream forms in the mammalian host.
Vertebrate
cristae
RQ* TCA cycle CN
sensitivity
Stumpy
+/-
0.12
- -
Slender
- 0.10
- -
Fly
Procyclic ++ 1.0
+ +
Epimastigote
+++
1.0
+ +
Metacyclic
+/-
0.1
- -
*RQ = respiratory
quotient (CO2 amount divided by O2 amount)
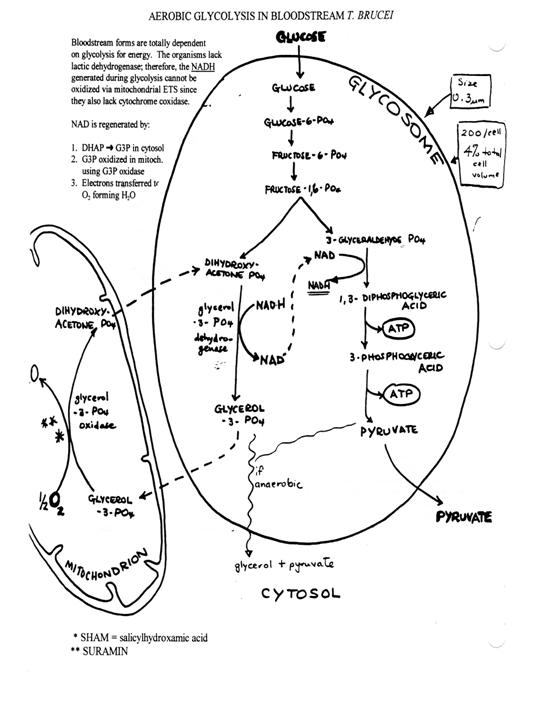

What is the basis
for the switch in metabolism? In the vertebrate
the parasite uses the regulatory mechanisms of the
host and utilizes the plentiful
energy source of the blood, glucose. The segregation of glycolytic enzymes in
the glycosome organelle substantially increases the efficiency of glycolysis.
Oxygen is consumed via a plant-like alternative oxidase, which does not produce
ATP by oxidative phosphorylation. The differentiation from the long slender to
the short stumpy form in the bloodstream involves changes in metabolism. The
stumpy forms are infective for the fly. In the fly
glucose is limiting and therefore a more efficient
utilization of glucose and amino acids occurs via
the TCA acid cycle and oxidative phosphorylation. Metacyclics
anticipate transfer to the vertebrate host by the
mitochondrion by losing cristae and TCA cycle
enzymes.
Differentiation of T. brucei
The differentiation from the stumpy bloodstream form into the procyclic form can
be studied in culture and involves massive mitochondrial biogenesis. It provides
a model system for the study of mitochondrial biogenesis in general.
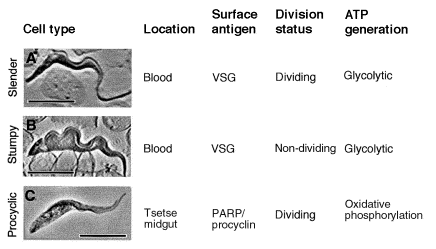
From
Mathews (1999). Long-slender
(A), short-stumpy (B) and procyclic forms (C) of the African trypanosome,
Trypanosoma brucei. The basic biological characteristics of each cell type
are shown to the right of each image. VSG, variable surface glycoprotein. Scale
bars = 10
 m.
m.
The differentiation from long slender (LS) to short stumpy (SS) occurs in
culture in so-called pleomorphic strains. It appears to be induced by cell
density apparently through a low molecular weight factor in the medium. Long
slenders are affected more by immune lysis than the short stumpies.

Also from
Mathews (1999). Representation of the different phases of the course of a
Trypanosoma brucei bloodstream parasitaemia. In Phase 1, the parasite
population increases in number due to proliferation of slender-form parasites
.Above a critical cell density, the slender-cell population initiates a phase of
`differentiation-divisions', which generate stumpy forms (Phase 2). The stumpy
forms do not divide and are competent for differentiation into the procyclic
form, either when taken up in a tsetse bloodmeal, or in vitro. In Phase
3, the population is composed predominantly of intermediate and stumpy-form
parasites; the population density eventually decreases as a result of
antibody-mediated clearance of first slender cells and then stumpy cells (Phase
4). Finally, the parasite population is re-established by the outgrowth of
slender-form parasites that have undergone antigenic switching (Phase 5).
The
differentiation from SS to procyclic cells can also be studied in vitro. It
is stimulated by the TCA cycle intermediate,
cisaconitate, and by lowering of the temperature to 27oC. Other
factors may also be involved in vivo such as glucose level and presence of
proteases. The process can be monitored by assaying the gain of the insect cell
-specific PARP cell surface protein and the loss of the bs-specific VSG coat.
AMINO ACIDS
AND PROTEINS
Little is known
about bloodstream forms. In T. gambiense the
major amino acid utilized is alanine, with glucose yielding
asp, glu, ala, gly. It also appears
that bloodstream forms can take up proteins. Culture forms use proline as an
energy source by reversal of the usual
biosynthetic pathway to
α- ketoglutarate.
Presumably all insect stages utilize proline. Trypanosomes lack catalase/peroxidase hemoproteins as well as
glutathione reductase. Thus they are sensitive to
nitrofurans (Nifurtimox) which produce high levels
of reactive oxygen intermediates—free radicals and
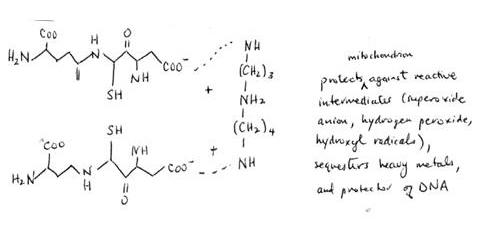
hydroxyl radicals. Parasites contain a unique reductant called trypanothionine, which consists of two glutathione peptides conjugated to spermidine. Reduction is via
trypanothionine reductase, an NADPH dependent
reaction.
Glutathione (glutamyl-cysteinyl-glycine)
+ spermidine = trypanothionine
Substrate
specificity min-1 M-1 GSH
reductase TSH reductase
Glutathione
1.8 x 108 0.8 x 108
Trypanothionine
1.4 x 108 5.0 x 108
Melarsoprol
inhibits trypanothionine reductase
Questions:
1. What is the difference between Salivarian and Stercorarian trypanosomes?
2. In the tsetse, the parasite undergoes a series of developmental changes. What are
these changes and where do they occur?
3. In the mammalian bloodstream, the parasites also undergo a developmental change.
What is the change and how is it stimulated?
4. What is the major difference in metabolism between the bloodstream forms and the
procyclic forms?
5. What is the alternate oxidase?